编者按:
为什么生活在高原的人,糖尿病发病率更低、血糖控制更好?这个问题已经困扰了医学界近百年。低温?紫外线?低氧?一直没人能说清楚。
最近,一项发表在顶级期刊Cell Metabolism上的研究终于揭开了谜底。答案不在肝脏、不在肌肉,而在身体里数量最多的细胞——红细胞里。这个发现,为未来糖尿病的治疗提供了一个全新的方向。

高原上血糖更好?一个困扰了医学界近百年的现象
故事缘起于20世纪20年代到40年代。
当时,哈佛大学的研究团队做了一次大胆的尝试:他们把一群健康志愿者送上了智利的安第斯山脉,海拔高达6000米。结果出乎所有人的意料——这些人的糖耐量竟然变好了。血糖更平稳,身体处理糖分的能力更强了。但在那个年代,没人能说清楚:为什么?
后来,陆续积累的证据表明,这并不是个例:
在海拔超过4000米的西藏人群中,糖尿病风险比低海拔地区降低89%;
在秘鲁4500米海拔生活的健康男性,空腹血糖比海平面居民低约15%;
在厄瓜多尔2700米左右海拔居住的大学生,高血糖风险降低75%。
更有趣的是,类似的现象不仅出现在人类身上,在高海拔的鹿鼠、藏猪、鸣禽中也同样存在。这说明:高原环境激活了一种非常古老、进化保守的降糖机制。
问题只剩下一个:高原上低温、紫外线、低氧……到底谁是真正的主角?
低氧,才是真正的“导演”
为了找出答案,研究人员把小鼠放进了模拟高海拔的低氧环境——8%的氧气,大约相当于5000米的海拔。结果令人惊讶:
小鼠血糖显著下降(图1A);
葡萄糖耐量明显改善(图1B);
效果在回到正常氧气后仍持续数周;
低氧小鼠的糖耐量虽然改善了,但胰岛素敏感性并未增强,反而有所下降(图D、E)。
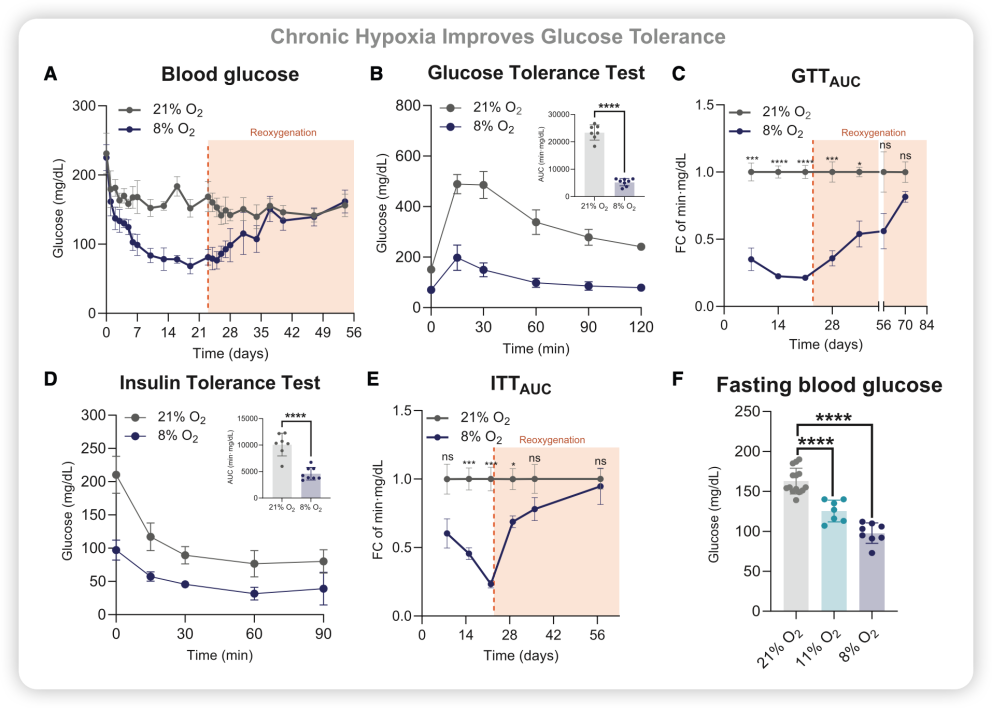
图1. 低氧对小鼠血糖稳态及糖耐量的影响
研究证实了低氧的降糖作用,更特别的是,这种降糖作用完全不依赖胰岛素。换句话说,身体走的是一条全新的“降糖通道”。
为了寻找这条通道,研究人员用PET/CT追踪葡萄糖的去向,发现了一个用传统降糖理论无法解释的现象:大脑、心脏、肝脏、肌肉……这些传统意义上的耗糖大户,加起来只能解释越30%的额外葡萄糖消耗(图2)。剩下约70%的糖,到底去了哪里?
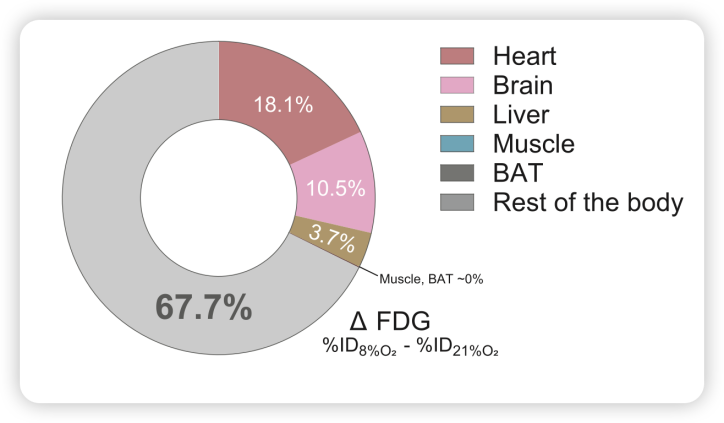
图2. 各器官对低氧下葡萄糖摄取增加的相对贡献
红细胞:被忽视的“葡萄糖黑洞”
研究人员把目光投向了一个长期被忽略的小角色——红细胞。
红细胞是人体内数量最多的细胞,成年人体内约85%的细胞是红细胞,但在代谢研究中,它们常常被忽略。
在血糖代谢方面,红细胞有一个非常特殊的地方:它们没有线粒体。线粒体是细胞的“发电厂”,能把葡萄糖高效地燃烧利用。没有线粒体,红细胞就只能靠另一种方式——糖酵解——来获取能量。糖酵解的效率很低,同样的葡萄糖只能产出极少能量。这意味着:红细胞必须不断“吞掉”大量的糖,才能维持正常生存。
换句话说,红细胞天生就是一个产能低下的“烧糖专业户”。
接下来的一系列实验,让研究人员确信:红细胞就是那个被忽视的“葡萄糖黑洞”。
1、证据一:减少红细胞 → 血糖回升
研究人员将低氧小鼠通过反复放血,把红细胞数量降到正常水平,它们的低血糖状态立即消失,糖耐量也回到不正常。
具体来看,研究人员每3天抽取小鼠15%的总血容量,连续4周。结果发现:放血后的低氧小鼠,红细胞压积降至与常氧对照组相当的水平,同时血浆葡萄糖从约130 mg/dl回升至约170 mg/dl,糖耐量曲线下的面积(AUC)也基本恢复到常氧水平(图3)。
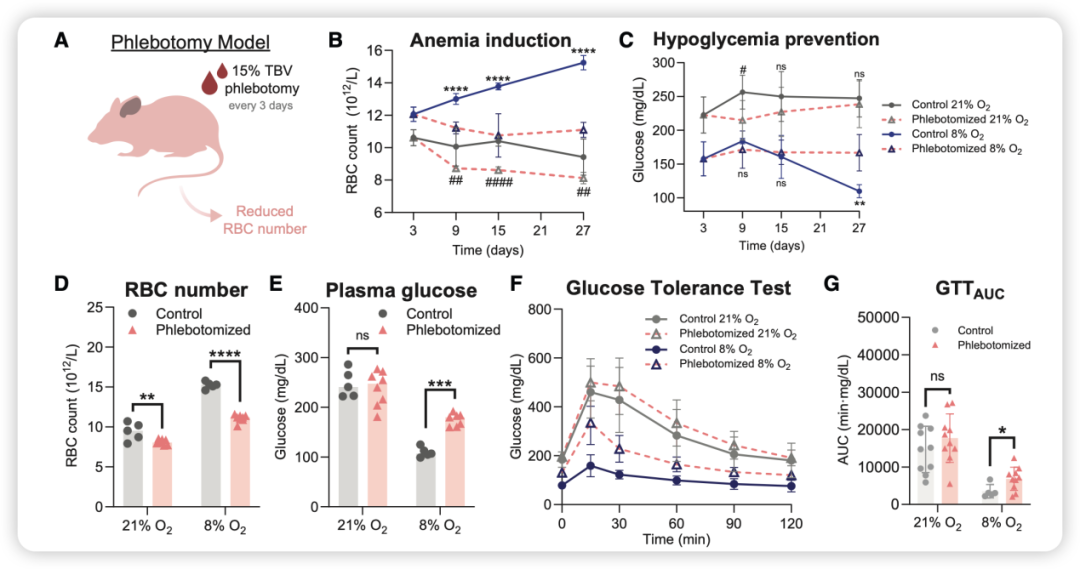
图3. 低氧对小鼠血糖稳态及糖耐量的影响
2、证据二:增加红细胞 → 血糖下降
为了进一步证实红细胞对血糖的影响,研究者进行了反向操作——在正常氧气环境下给小鼠输注浓缩红细胞(75%压积,连续两天,每天两次),结果发现:
无论输注的是来自常氧供体还是低氧供体的红细胞,受体小鼠的红细胞数量都增加了约20%~30%,血糖随之下降约20~30 mg/dl。其中,接受低氧供体红细胞的组别降糖效果略强,提示低氧红细胞本身还具有更强的“质”的优势(图4)。
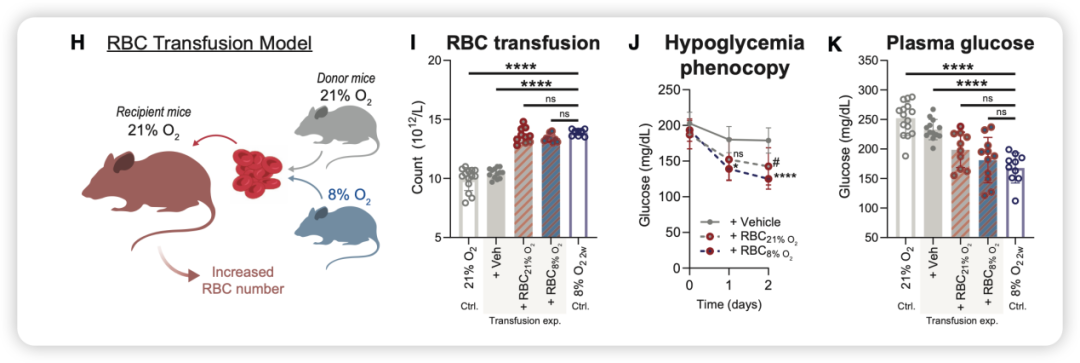
图4. 红细胞输注实验——红细胞数量增加足以降低血糖
3、证据三:低氧让单个红细胞“更吃糖”
在充分证实红细胞对血糖代谢的影响之后,研究者还需要确认低氧除了增加了红细胞的数量,能否增强单个红细胞的摄糖功能。
研究人员用稳定同位素标记的2-脱氧葡萄糖(2DG-U-13C)进行体内示踪,发现低氧小鼠红细胞内磷酸化2DG的累积速度是对照组的3倍。在离体共孵育实验中,将低氧来源的红细胞与等量血浆混合(初始葡萄糖浓度相同),10分钟后测量剩余葡萄糖,计算出单个红细胞的葡萄糖摄取能力提高了2.5倍。
而且,低氧只能影响新生红细胞的摄糖能力。研究人员用生物素标记追踪发现:低氧环境下新生成的红细胞,其GLUT1葡萄糖转运蛋白表达量比老细胞高出60%,GLUT4也上调约48%。而老细胞(生物素阳性)的GLUT1水平在常氧和低氧之间没有显著差异。
以上系列试验证实了低氧通过两条途径重塑红细胞的降糖功能:一是数量扩增——红细胞总数增加,直接扩大葡萄糖消耗总量;二是质量提升——提高新生红细胞的摄糖能力。这完美地解释了为什么高原上血糖更好,困扰医学界近百年的谜团彻底揭开。
答案藏在红细胞膜上一个叫 band 3的蛋白身上。
常氧时:GAPDH(糖酵解关键酶)被band 3粘在细胞膜上,糖酵解被抑制。
低氧时:脱氧血红蛋白与band 3结合,把GAPDH挤下来,释放到细胞浆中,糖酵解被激活。
研究人员用多种技术验证了这一机制:免疫荧光和STED超分辨显微镜显示GAPDH从膜上转移到胞浆;邻位连接技术证明GAPDH?band3相互作用减少约50%~70%;交联质谱则精确找到了脱氧血红蛋白“挤走”GAPDH的结合位点。
这个机制在小鼠和人类红细胞中高度保守,而且响应速度极快——几分钟内就能完成。
红细胞,糖尿病治疗的新希望
最重要的部分来了:这个机制能不能用来治疗糖尿病?答案是:可以。
1、1型糖尿病模型(STZ诱导)
研究人员用链脲佐菌素(STZ)连续5天注射小鼠,摧毁胰岛β细胞,建立1型糖尿病模型。待血糖稳定升高后(约2周),将小鼠随机分配到常氧(21% O2)或低氧(8% O2)环境中,持续3周。结果显示(图5):
低氧治疗组的糖尿病小鼠,血糖从初始的约400~500 mg/dl降至接近200 mg/dl,降幅超过50%;
葡萄糖耐量测试显示,低氧组曲线下面积(AUC)几乎恢复到正常小鼠水平;
即使不改变环境氧气,仅输注低氧供体的红细胞(连续2天,每天2次),也能使STZ小鼠的血糖在2天内下降约100~150 mg/dl。
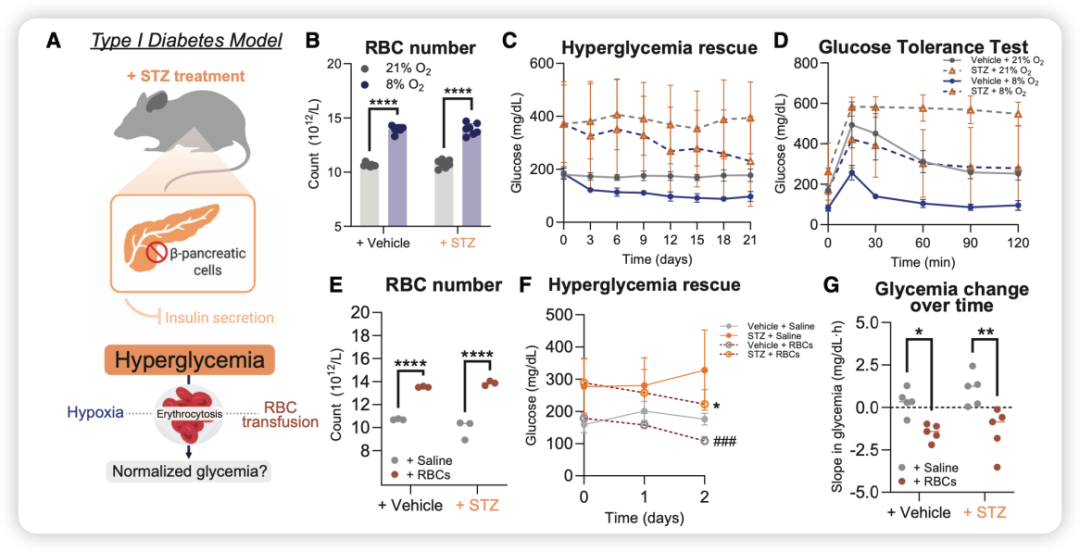
图5. 低氧与红细胞输注对1型糖尿病模型的血糖挽救效果
研究人员使用了他们自主研发的小分子药物——HypoxyStat。该药物能结合血红蛋白,增加其氧亲和力,从而限制氧在组织中的释放,在常氧环境下模拟出组织缺氧的状态。
实验设计:小鼠分别喂食高脂饮食(HFD)或普通饮食(CD),持续数周诱导肥胖和胰岛素抵抗;随后每天口服HypoxyStat(600?mg/kg)或溶剂对照,持续2.5周,全程在常氧环境中饲养。结果非常明确(图6):
高脂饮食+HypoxyStat组小鼠的空腹血糖完全恢复正常,从治疗前的约200~250 mg/dl降至140~160 mg/dl;
该组小鼠的糖耐量也恢复到与正常饮食组无显著差异的水平;
治疗组小鼠红细胞计数增加约30%~40%,且未对其他血常规指标产生不良影响;
更重要的是,HypoxyStat的效果是可逆的——停药后红细胞计数和血糖水平会逐渐恢复至基线。
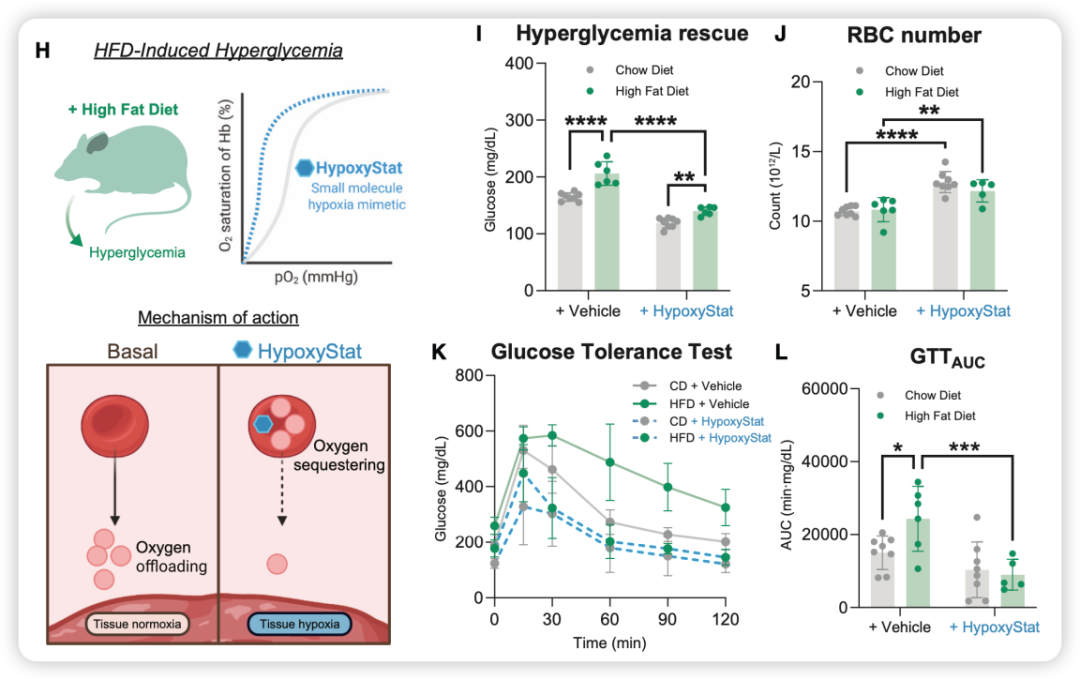
图6. 小分子低氧模拟物HypoxyStat挽救高脂饮食诱导的2型糖尿病高血糖
无论是低氧、还是口服HypoxyStat,都能通过激活红细胞的降糖能力,在不依赖胰岛素的情况下,逆转1型和2型糖尿病小鼠的高血糖。这项研究为糖尿病治疗带来了真正的新希望。
研究局限及未来展望
诚然,这项研究听起来令人振奋。但也有人会问:这是否意味着该成果可以立即应用于临床患者?答案是否定的。本研究虽然开辟了全新的研究方向,但距离真正的临床转化,仍有一段距离。
首先,实验目前仅在雄性小鼠中进行。红细胞生成和代谢在雌性、老年或其他品系小鼠中是否存在差异,还需要进一步验证。进一步在人体进行研究,还有漫长的道路要走。
其次,长期提高红细胞压积,可能会增加血液黏稠度,带来健康风险。因此,在转化为人体治疗时,需要谨慎评估风险与收益。
一个潜在的解决思路是:促进红细胞更新,让更多年轻、高葡萄糖摄取能力的红细胞进入循环,而不是简单地增加红细胞总数。这样既可以利用新生红细胞的降糖优势,又能避免血液黏稠度过高带来的心血管负担。
无论如何,这项研究为糖尿病治疗打开了一个全新的方向:从盯着胰腺、肝脏、肌肉,到关注红细胞——代谢性疾病治疗的地图,又多了一块重要拼图。
参考文献:Marti-Mateos Y, Safari Z, Bevers S, et al. Red blood cells serve as a primary glucose sink to improve glucose tolerance at altitude. *Cell Metabolism*. 2026;38(3):529-545.e8. 2 comments
为什么生活在高原的人,糖尿病发病率更低、血糖控制更好?这个问题已经困扰了医学界近百年。低温?紫外线?低氧?一直没人能说清楚。
最近,一项发表在顶级期刊Cell Metabolism上的研究终于揭开了谜底。答案不在肝脏、不在肌肉,而在身体里数量最多的细胞——红细胞里。这个发现,为未来糖尿病的治疗提供了一个全新的方向。
高原上血糖更好?一个困扰了医学界近百年的现象
故事缘起于20世纪20年代到40年代。
当时,哈佛大学的研究团队做了一次大胆的尝试:他们把一群健康志愿者送上了智利的安第斯山脉,海拔高达6000米。结果出乎所有人的意料——这些人的糖耐量竟然变好了。血糖更平稳,身体处理糖分的能力更强了。但在那个年代,没人能说清楚:为什么?
后来,陆续积累的证据表明,这并不是个例:
在海拔超过4000米的西藏人群中,糖尿病风险比低海拔地区降低89%;
在秘鲁4500米海拔生活的健康男性,空腹血糖比海平面居民低约15%;
在厄瓜多尔2700米左右海拔居住的大学生,高血糖风险降低75%。
更有趣的是,类似的现象不仅出现在人类身上,在高海拔的鹿鼠、藏猪、鸣禽中也同样存在。这说明:高原环境激活了一种非常古老、进化保守的降糖机制。
问题只剩下一个:高原上低温、紫外线、低氧……到底谁是真正的主角?
低氧,才是真正的“导演”
为了找出答案,研究人员把小鼠放进了模拟高海拔的低氧环境——8%的氧气,大约相当于5000米的海拔。结果令人惊讶:
小鼠血糖显著下降(图1A);
葡萄糖耐量明显改善(图1B);
效果在回到正常氧气后仍持续数周;
低氧小鼠的糖耐量虽然改善了,但胰岛素敏感性并未增强,反而有所下降(图D、E)。
图1. 低氧对小鼠血糖稳态及糖耐量的影响
研究证实了低氧的降糖作用,更特别的是,这种降糖作用完全不依赖胰岛素。换句话说,身体走的是一条全新的“降糖通道”。
为了寻找这条通道,研究人员用PET/CT追踪葡萄糖的去向,发现了一个用传统降糖理论无法解释的现象:大脑、心脏、肝脏、肌肉……这些传统意义上的耗糖大户,加起来只能解释越30%的额外葡萄糖消耗(图2)。剩下约70%的糖,到底去了哪里?
图2. 各器官对低氧下葡萄糖摄取增加的相对贡献
红细胞:被忽视的“葡萄糖黑洞”
研究人员把目光投向了一个长期被忽略的小角色——红细胞。
红细胞是人体内数量最多的细胞,成年人体内约85%的细胞是红细胞,但在代谢研究中,它们常常被忽略。
在血糖代谢方面,红细胞有一个非常特殊的地方:它们没有线粒体。线粒体是细胞的“发电厂”,能把葡萄糖高效地燃烧利用。没有线粒体,红细胞就只能靠另一种方式——糖酵解——来获取能量。糖酵解的效率很低,同样的葡萄糖只能产出极少能量。这意味着:红细胞必须不断“吞掉”大量的糖,才能维持正常生存。
换句话说,红细胞天生就是一个产能低下的“烧糖专业户”。
接下来的一系列实验,让研究人员确信:红细胞就是那个被忽视的“葡萄糖黑洞”。
1、证据一:减少红细胞 → 血糖回升
研究人员将低氧小鼠通过反复放血,把红细胞数量降到正常水平,它们的低血糖状态立即消失,糖耐量也回到不正常。
具体来看,研究人员每3天抽取小鼠15%的总血容量,连续4周。结果发现:放血后的低氧小鼠,红细胞压积降至与常氧对照组相当的水平,同时血浆葡萄糖从约130 mg/dl回升至约170 mg/dl,糖耐量曲线下的面积(AUC)也基本恢复到常氧水平(图3)。
图3. 低氧对小鼠血糖稳态及糖耐量的影响
2、证据二:增加红细胞 → 血糖下降
为了进一步证实红细胞对血糖的影响,研究者进行了反向操作——在正常氧气环境下给小鼠输注浓缩红细胞(75%压积,连续两天,每天两次),结果发现:
无论输注的是来自常氧供体还是低氧供体的红细胞,受体小鼠的红细胞数量都增加了约20%~30%,血糖随之下降约20~30 mg/dl。其中,接受低氧供体红细胞的组别降糖效果略强,提示低氧红细胞本身还具有更强的“质”的优势(图4)。
图4. 红细胞输注实验——红细胞数量增加足以降低血糖
3、证据三:低氧让单个红细胞“更吃糖”
在充分证实红细胞对血糖代谢的影响之后,研究者还需要确认低氧除了增加了红细胞的数量,能否增强单个红细胞的摄糖功能。
研究人员用稳定同位素标记的2-脱氧葡萄糖(2DG-U-13C)进行体内示踪,发现低氧小鼠红细胞内磷酸化2DG的累积速度是对照组的3倍。在离体共孵育实验中,将低氧来源的红细胞与等量血浆混合(初始葡萄糖浓度相同),10分钟后测量剩余葡萄糖,计算出单个红细胞的葡萄糖摄取能力提高了2.5倍。
而且,低氧只能影响新生红细胞的摄糖能力。研究人员用生物素标记追踪发现:低氧环境下新生成的红细胞,其GLUT1葡萄糖转运蛋白表达量比老细胞高出60%,GLUT4也上调约48%。而老细胞(生物素阳性)的GLUT1水平在常氧和低氧之间没有显著差异。
以上系列试验证实了低氧通过两条途径重塑红细胞的降糖功能:一是数量扩增——红细胞总数增加,直接扩大葡萄糖消耗总量;二是质量提升——提高新生红细胞的摄糖能力。这完美地解释了为什么高原上血糖更好,困扰医学界近百年的谜团彻底揭开。
开关机制:一个精妙的“氧传感器”
答案藏在红细胞膜上一个叫 band 3的蛋白身上。
常氧时:GAPDH(糖酵解关键酶)被band 3粘在细胞膜上,糖酵解被抑制。
低氧时:脱氧血红蛋白与band 3结合,把GAPDH挤下来,释放到细胞浆中,糖酵解被激活。
研究人员用多种技术验证了这一机制:免疫荧光和STED超分辨显微镜显示GAPDH从膜上转移到胞浆;邻位连接技术证明GAPDH?band3相互作用减少约50%~70%;交联质谱则精确找到了脱氧血红蛋白“挤走”GAPDH的结合位点。
这个机制在小鼠和人类红细胞中高度保守,而且响应速度极快——几分钟内就能完成。
红细胞,糖尿病治疗的新希望
最重要的部分来了:这个机制能不能用来治疗糖尿病?答案是:可以。
1、1型糖尿病模型(STZ诱导)
研究人员用链脲佐菌素(STZ)连续5天注射小鼠,摧毁胰岛β细胞,建立1型糖尿病模型。待血糖稳定升高后(约2周),将小鼠随机分配到常氧(21% O2)或低氧(8% O2)环境中,持续3周。结果显示(图5):
低氧治疗组的糖尿病小鼠,血糖从初始的约400~500 mg/dl降至接近200 mg/dl,降幅超过50%;
葡萄糖耐量测试显示,低氧组曲线下面积(AUC)几乎恢复到正常小鼠水平;
即使不改变环境氧气,仅输注低氧供体的红细胞(连续2天,每天2次),也能使STZ小鼠的血糖在2天内下降约100~150 mg/dl。
图5. 低氧与红细胞输注对1型糖尿病模型的血糖挽救效果
2、2型糖尿病模型(高脂饮食诱导)
研究人员使用了他们自主研发的小分子药物——HypoxyStat。该药物能结合血红蛋白,增加其氧亲和力,从而限制氧在组织中的释放,在常氧环境下模拟出组织缺氧的状态。
实验设计:小鼠分别喂食高脂饮食(HFD)或普通饮食(CD),持续数周诱导肥胖和胰岛素抵抗;随后每天口服HypoxyStat(600?mg/kg)或溶剂对照,持续2.5周,全程在常氧环境中饲养。结果非常明确(图6):
高脂饮食+HypoxyStat组小鼠的空腹血糖完全恢复正常,从治疗前的约200~250 mg/dl降至140~160 mg/dl;
该组小鼠的糖耐量也恢复到与正常饮食组无显著差异的水平;
治疗组小鼠红细胞计数增加约30%~40%,且未对其他血常规指标产生不良影响;
更重要的是,HypoxyStat的效果是可逆的——停药后红细胞计数和血糖水平会逐渐恢复至基线。
图6. 小分子低氧模拟物HypoxyStat挽救高脂饮食诱导的2型糖尿病高血糖
无论是低氧、还是口服HypoxyStat,都能通过激活红细胞的降糖能力,在不依赖胰岛素的情况下,逆转1型和2型糖尿病小鼠的高血糖。这项研究为糖尿病治疗带来了真正的新希望。
研究局限及未来展望
诚然,这项研究听起来令人振奋。但也有人会问:这是否意味着该成果可以立即应用于临床患者?答案是否定的。本研究虽然开辟了全新的研究方向,但距离真正的临床转化,仍有一段距离。
首先,实验目前仅在雄性小鼠中进行。红细胞生成和代谢在雌性、老年或其他品系小鼠中是否存在差异,还需要进一步验证。进一步在人体进行研究,还有漫长的道路要走。
其次,长期提高红细胞压积,可能会增加血液黏稠度,带来健康风险。因此,在转化为人体治疗时,需要谨慎评估风险与收益。
一个潜在的解决思路是:促进红细胞更新,让更多年轻、高葡萄糖摄取能力的红细胞进入循环,而不是简单地增加红细胞总数。这样既可以利用新生红细胞的降糖优势,又能避免血液黏稠度过高带来的心血管负担。
无论如何,这项研究为糖尿病治疗打开了一个全新的方向:从盯着胰腺、肝脏、肌肉,到关注红细胞——代谢性疾病治疗的地图,又多了一块重要拼图。
参考文献:Marti-Mateos Y, Safari Z, Bevers S, et al. Red blood cells serve as a primary glucose sink to improve glucose tolerance at altitude. *Cell Metabolism*. 2026;38(3):529-545.e8. 2 comments








 京公网安备 11010502033361号
京公网安备 11010502033361号
发布留言