一、T1DM流行病学:全球攀升的疾病负担与中国特色挑战
全球疾病负担沉重
T1DM以胰岛β细胞进行性破坏为核心,并发症风险高。糖尿病控制与并发症试验/糖尿病干预与并发症流行病学研究(DCCT/EDIC)显示[1],患者30年累积发病率:增殖性糖尿病视网膜病变(PDR)33.4%、临床显著性黄斑水肿36.6%、估算肾小球滤过率(eGFR)降低14.1%、截肢3.2%、心血管疾病(CVD)17.1%、死亡率10.4%(图1),疾病负担随病程显著加重。
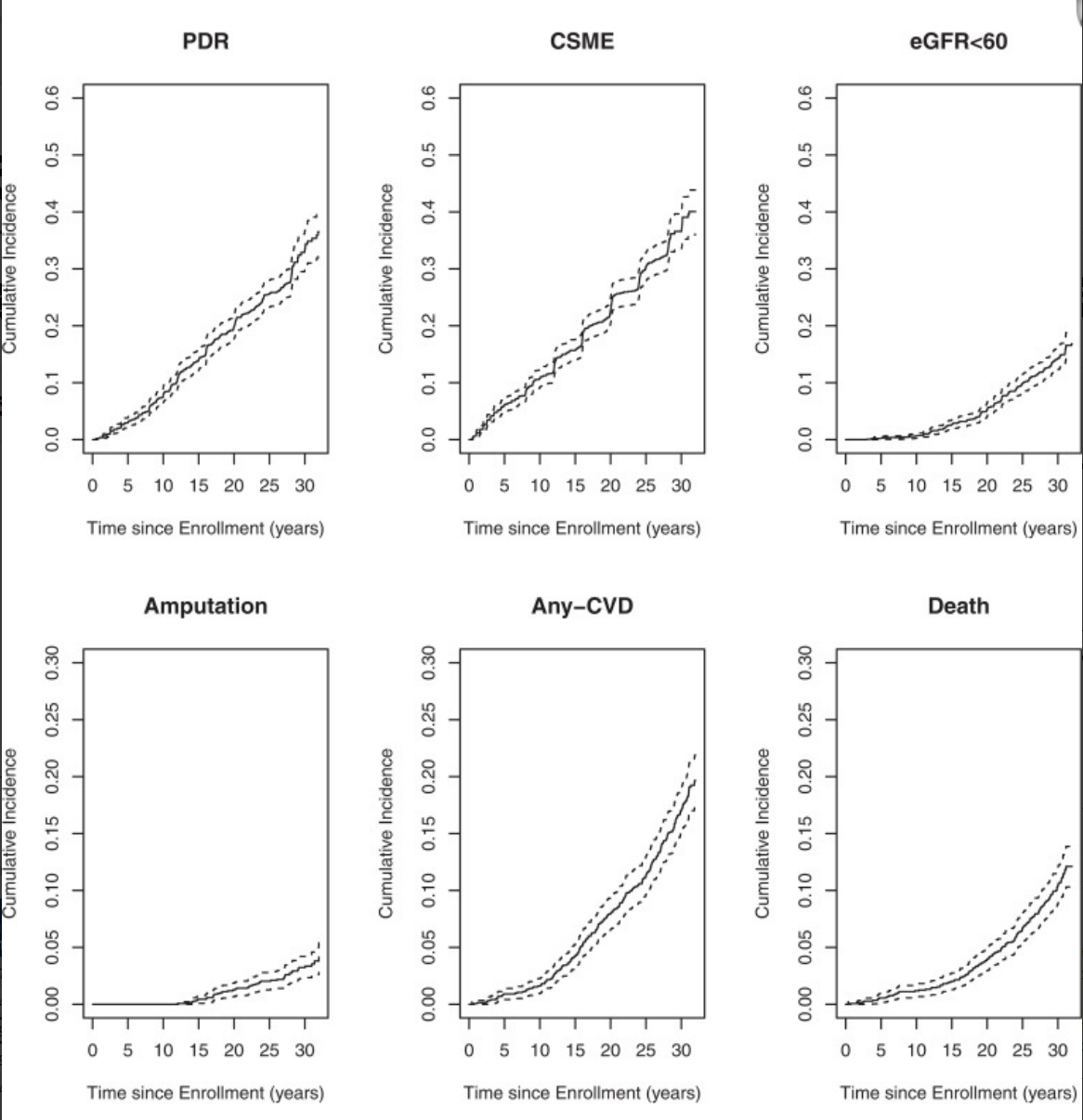
图1. T1DM患者30年并发症累积发病率曲线
中国T1DM:高患病人数与LADA漏诊
2024年全球T1DM患病人数排名中,中国以59.9万所有年龄段患者、11.7万儿童及青少年(<20岁)患者居第三。更突出的是成人隐匿性自身免疫糖尿病(LADA)漏诊问题:在年龄≥30岁、谷氨酸脱羧酶抗体(GADA)阳性的新诊断表型2型糖尿病(T2DM)中,LADA占比5.9%;在新诊断表型T1DM中,LADA占比65%,且高GADA滴度LADA占比达1/2,因症状与T2DM相似易漏诊。
发病核心:遗传、环境与免疫因素交互损伤胰岛β细胞
遗传风险基因&免疫:T1DM的风险基因编码HLA分子、共刺激/共抑制蛋白、受体/配体、细胞因子及其受体等,参与T细胞激活/分化,影响T细胞发育和功能[2]。关键HLA-Ⅱ类基因(如HLA-DR4、HLA-DQ8等)突变导致自身抗原呈递异常,是T1DM最强遗传风险因素,与非HLA基因协同打破免疫稳态(图2)。
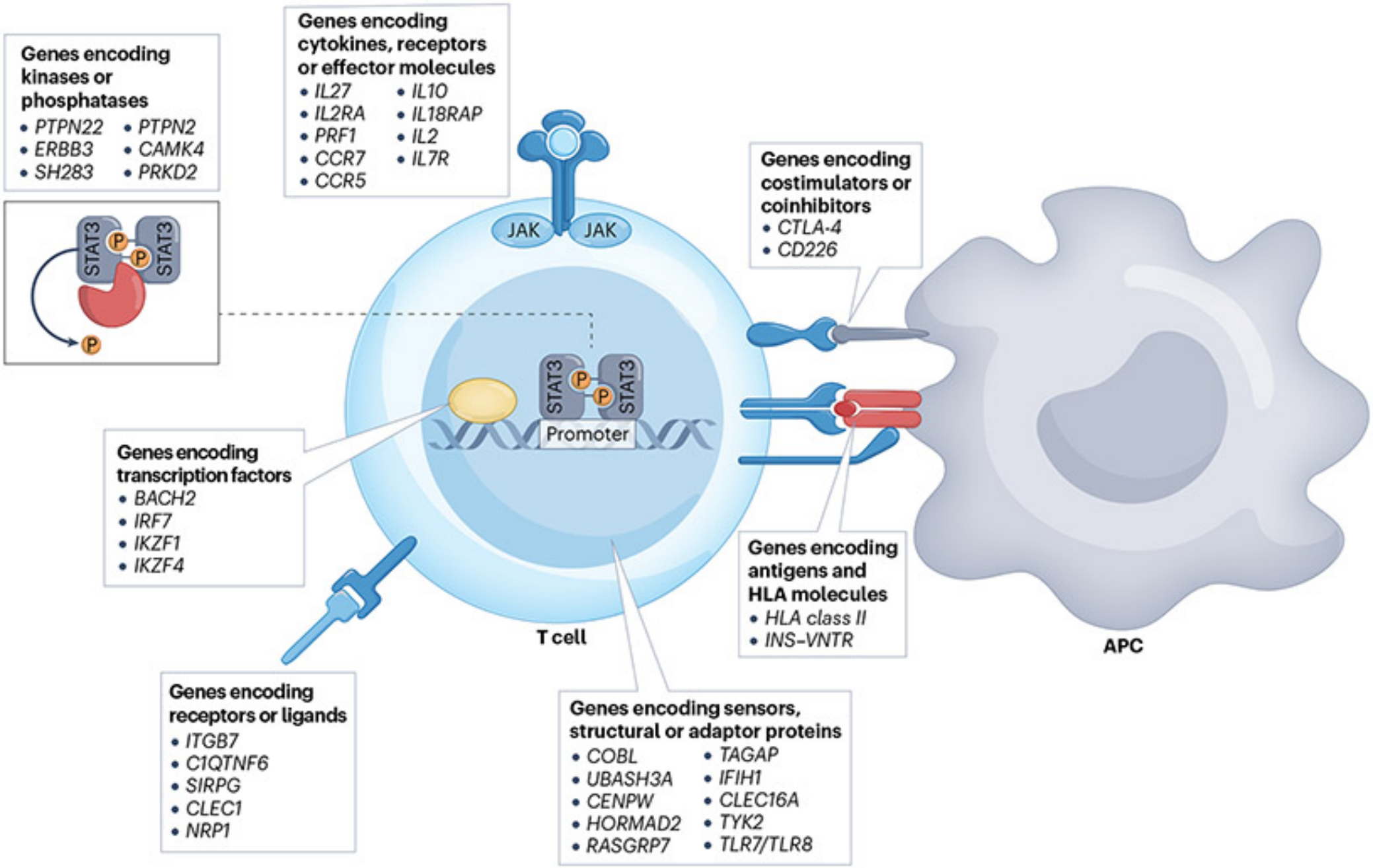
图2. T1DM风险基因介导的抗原呈递与T细胞活化机制
环境因素&免疫:环境因素可通过调节免疫细胞分化改变T细胞亚群平衡、诱导β细胞表达抗原,促进自身免疫应答的发生和发展[2]。具体来说,环境因素可促进CD4+T细胞分化、CD8+T细胞及B细胞活化,攻击胰岛β细胞;抑制Treg细胞,降低免疫耐受;激活抗原呈递细胞(APC),放大T细胞应答;环境因素/细胞因子可诱发β细胞内质网应激,上调HLA或PD-L1表达,使β细胞被免疫识别攻击(图3)。
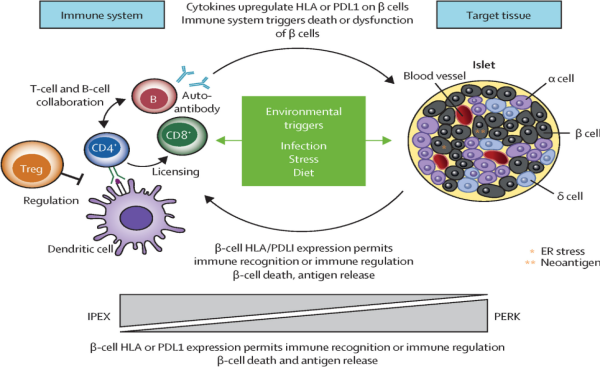
图3. 环境与免疫驱动T1DM的β细胞损伤机制
肠道微生物是关键环境因素:肠道微生物失调可能通过破坏肠屏障、调节肠道免疫应答及丧失共生菌耐受等途径驱动疾病发生发展。菌群多样性降低及代谢产物短链脂肪酸(SCFAs)缺乏,会增加肠壁通透性,导致食物或菌群抗原入血,激活自身反应性T细胞,影响肠道黏膜稳定性T细胞功能,引发局部免疫失衡,误杀共生菌,产生交叉反应性抗体/T细胞,促使致病菌增殖,加剧炎症反应,进而影响整体免疫应答[2]。
二、T1DM肠道微生物巨变
动物模型研究:菌群移植可调控T1DM发病进程
NOD小鼠研究:将新发T1DM儿童粪菌移植给无菌非肥胖糖尿病(NOD)小鼠,其菌群α多样性较供体显著降低,且T1DM发病延迟,证实菌群结构直接影响疾病进程(图4)[3]。
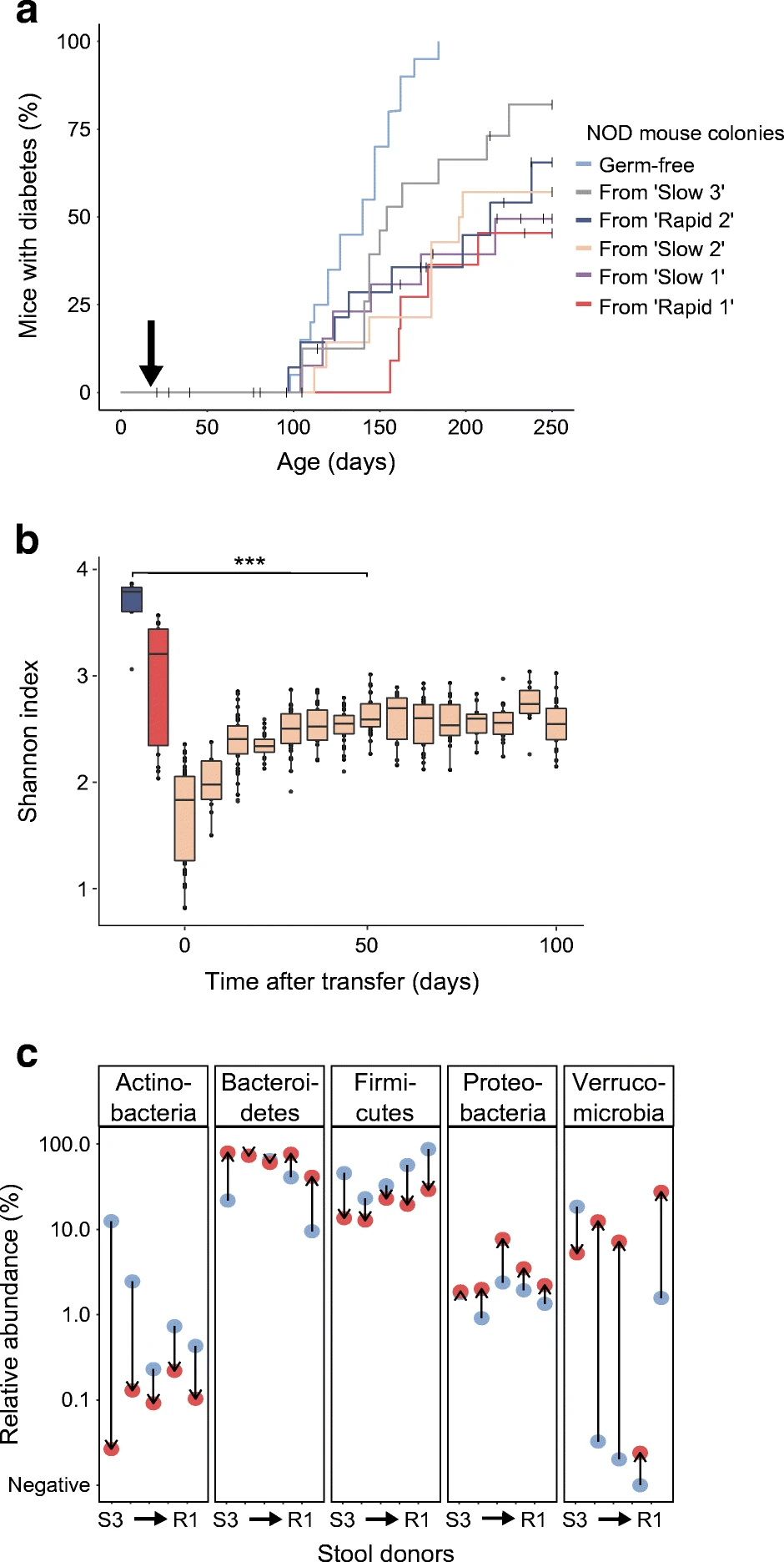
图4. 肠道菌群移植后细菌定植与生存分析
Akk菌的保护作用:链脲佐菌素(STZ)造模小鼠中,Akkermansia muciniphila丰度呈“先降后升”动态变化;外源移植该菌可促进肠道黏液生成,上调抗菌肽Reg3γ表达,降低胰岛Toll样受体4(TLR4)水平,使Treg细胞比例升高23%,显著延缓糖尿病发生(图5)[4]。
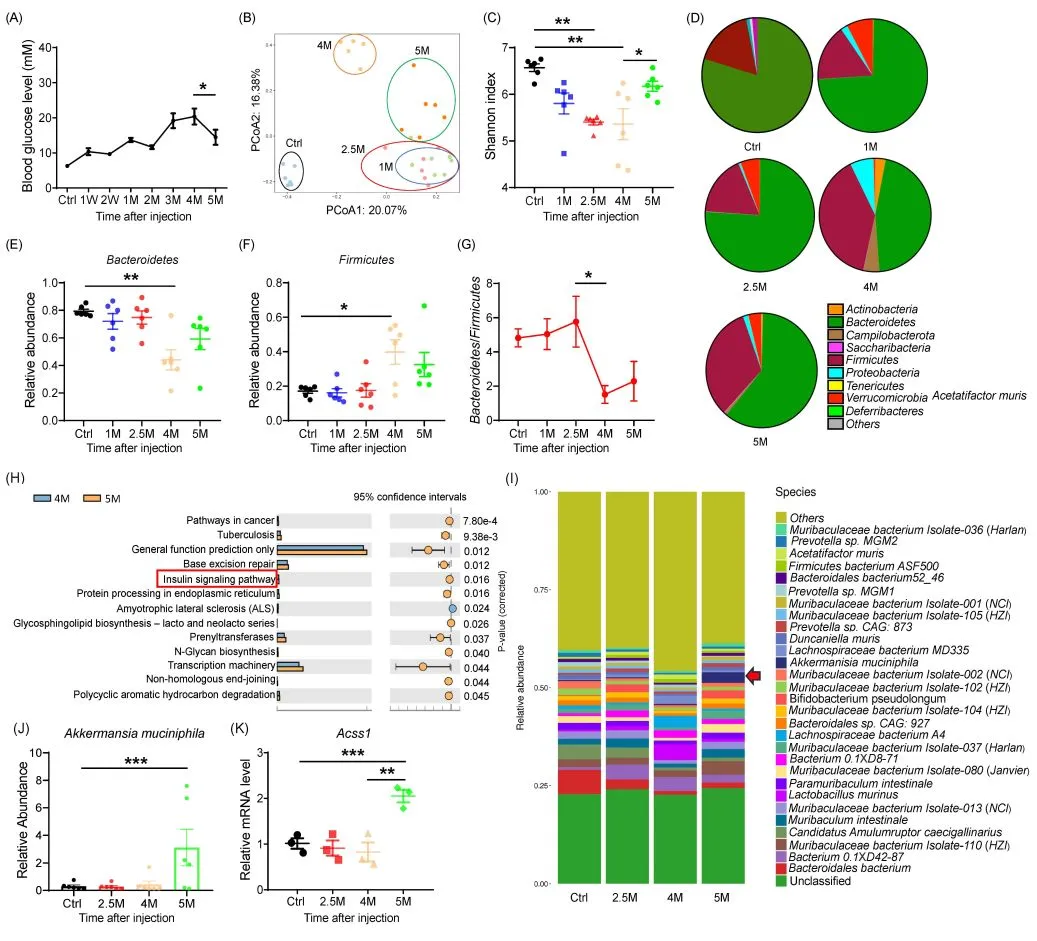
图5. T1DM小鼠相关肠道菌群动态变化特征
临床人群研究:T1DM患者菌群与代谢物的特征性紊乱
T1DM患儿肠道菌群呈现“有益菌减少、致病菌增加”的失衡态:厚壁菌门产丁酸盐菌(粪杆菌属、瘤胃球菌属-2)丰度较健康儿童降低40%,拟杆菌属、志贺氏埃希氏菌属升高2倍以上[5,6]。
成人亚型对比研究显示,LADA患者SCFAs菌丰度较GADA阴性T1DM患者低17%,且血清脯氨酸衍生物水平升高32%,而SCFAs可通过激活GPR43受体抑制Th17细胞分化。这一研究证实,GADA是影响成人T1DM肠道菌群结构的重要因素,且T1DM自身免疫状态与产SCFAs菌缺乏密切相关[7]。
菌群-代谢物-免疫的调控网络:研究证实,粪杆菌属与促炎因子IL-6、胰岛自身抗体GADA均呈负相关;粪便支链氨基酸/芳香族氨基酸比值与胰岛功能指数(HOMA-β)呈正相关,提示菌群代谢物可直接调节免疫应答与胰岛功能[7]。
三、T1DM胰腺免疫狙击战:菌群驱动的免疫损伤
“肠道菌群失衡相关性自身免疫综合征”理论
失调菌群通过三重途径触发自身免疫:①破坏肠屏障使脂多糖(LPS)入血,激活TLR4信号通路;②致病菌抗原与胰岛抗原分子模拟,引发交叉免疫反应;③抑制Treg细胞分化,打破免疫耐受。这一机制在系统性红斑狼疮、弥漫性毒性甲状腺肿等疾病中均有印证,提示肠道菌群是自身免疫疾病的共同驱动因素。
T1DM患者的免疫特征
T1DM患者存在显著的肠道菌群巨变,核心特征是产SCFAs菌丰度降低,且胰岛自身抗体阳性T1DM患者的菌群失调程度比阴性患者更显著。实验证据显示,Akk菌可改善STZ模型小鼠的高血糖,灌胃灭活Akk菌能提升菌群多样性、激活肠免疫通路;抗菌肽(CRAMP)则可预防NOD小鼠患T1DM——NOD母鼠口服表达CRAMP的益生菌后,子鼠菌群持久改善,异常干扰素信号消失,T1DM发病率下降(图6)[8]。
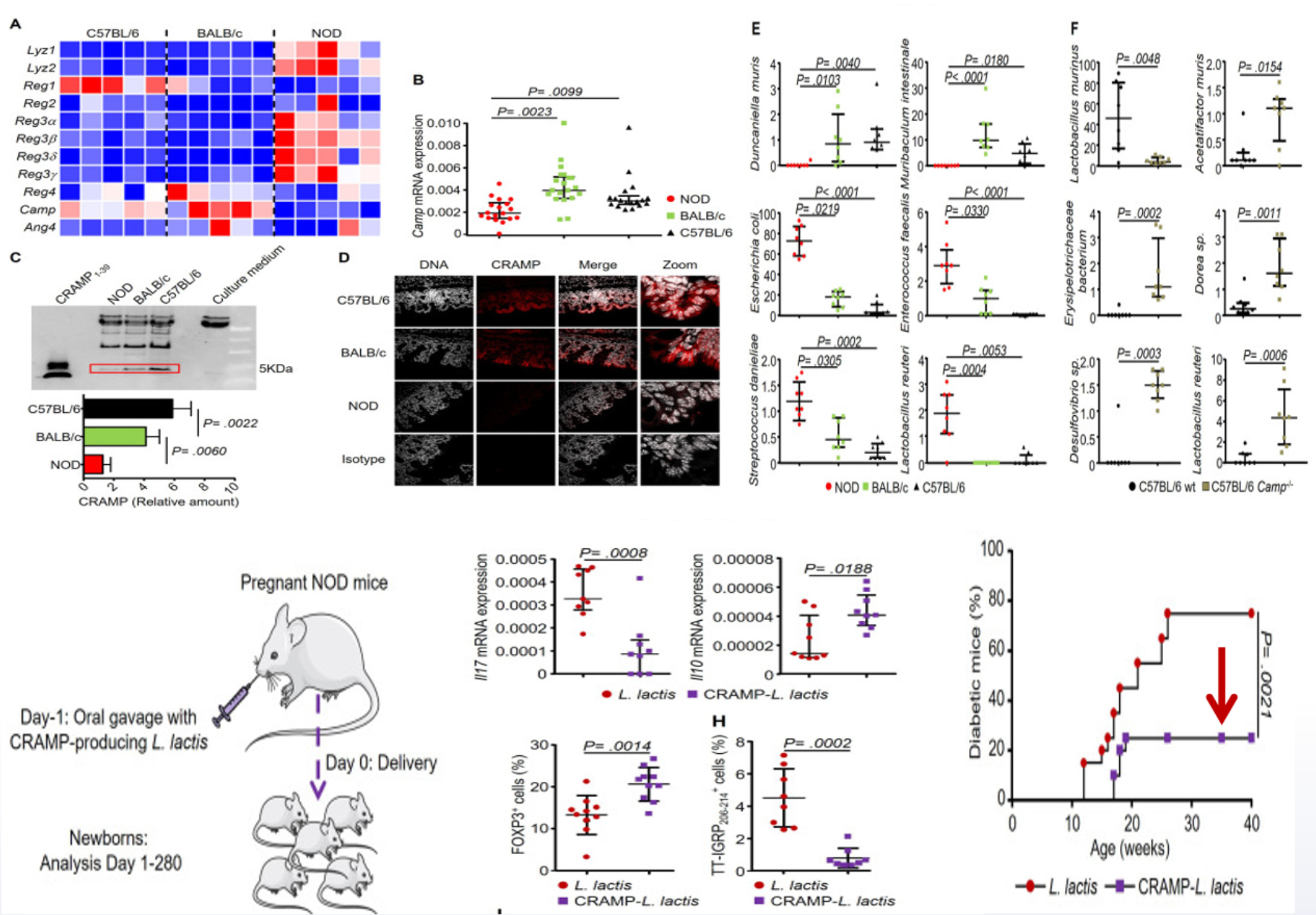
图6. CRAMP预防NOD小鼠患T1DM的机制研究
在袁慧娟教授团队近期研究中,通过流式细胞术检测发现,无论T1DM患者是否为GADA阳性,其外周血T细胞亚群失衡模式一致:CD4+/CD8+T细胞、Th1/Th17/Tc1/Tc17细胞增多,Treg、Th2/Tc2细胞减少,提示细胞免疫损伤胰腺β细胞的过程无显著差异。进一步研究表明,产SCFAs菌与Treg正相关、与CD8+T细胞负相关,特定菌群可通过调控树突状细胞功能或TLR通路,影响T细胞分化。
粪菌移植的干预证据
袁慧娟教授团队通过粪菌移植实验进一步验证了菌群的作用:将T1DM患者粪菌移植给免疫正常的C57小鼠,仅导致肠屏障受损,无血糖升高或胰岛损伤;而移植给免疫缺陷的NOD小鼠后,小鼠血糖显著升高、胰岛素分泌减少,胰腺出现淋巴细胞浸润,粪便SCFAs含量降低,胰腺T细胞亚群紊乱(CD4+/CD8+T细胞增多、Treg减少)。综上,T1DM失调菌群无法单独致病,需在遗传或免疫缺陷背景下才会“狙击”胰腺;无论T1DM患者是否合并胰岛自身抗体阳性,胰腺细胞免疫损伤机制一致,仅T1DM患者菌群失调与免疫紊乱更严重。这一研究为T1DM的预防与治疗提供了“菌群调节+免疫修复”的潜在方向。
四、结语
袁慧娟教授的报告系统论证了肠道菌群在T1DM发病中的核心驱动作用。研究结论明确指出,T1DM的本质是遗传背景下,由菌群失调触发的免疫耐受失衡。其核心机制在于:产SCFAs菌的减少破坏了肠屏障与免疫稳态,导致Treg细胞缺失与效应T细胞(CD4+/CD8+)的过度活化,最终引发对胰岛β细胞的免疫攻击。值得注意的是,无论患者GADA抗体是否阳性,其胰腺免疫损伤的细胞学机制一致,仅严重程度有别。这一发现将T1DM的病因聚焦于“菌群-免疫”轴,为临床探索“菌群调节+免疫修复”的整合治疗策略奠定了坚实的理论基础。
参考文献
1.Bebu I, et al. Diabetes Care. 2023;46(2):361-368.
2.Nat Rev Endocrinol 2024.
3.Crèvecoeur I, et al. Diabetologia. 2017;60(3):475-489.
4.Zhang C, et al. Imeta. 2024 Oct 1;3(5):e237.
5.Kostic AD, et al. Cell Host Microbe. 2015;17(2):260-273.
6.Yuan X, et al. Nat Commun. 2022;13(1):6356.
7.Fang Y, et al. Diabetes Care. 2021;44(12):2738-2746.
8.Liang W, et al. Gastroenterology. 2022;162(4):1288-1302.e16.
2 comments








 京公网安备 11010502033361号
京公网安备 11010502033361号
发布留言